Conradie, Bruggeman, Ciliberto, Csikasz-Nagy, Novak, Westerhoff, Snoep,
Model Status
This model has been built using the expressions found in the Conradie's 2010 paper "Restriction point control of the mammalian cell cycle via the cyclin E/Cdk2:p27 complex". This model was produced using the parameter values on page 360. This file is known to run on COR 0.9 and Open Cell.
Model Structure
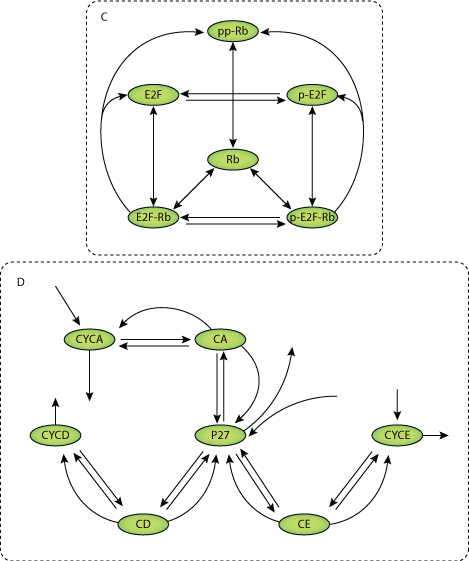
Numerous top-down kinetic models have been constructed to describe the cell cycle. These models have typically been constructed, validated and analyzed using model species (molecular intermediates and proteins) and phenotypic observations, and therefore do not focus on the individual model processes (reaction steps). We have developed a method to: (a) quantify the importance of each of the reaction steps in a kinetic model for the positioning of a switch point [i.e. the restriction point (RP)]; (b) relate this control of reaction steps to their effects on molecular species, using sensitivity and co-control analysis; and thereby (c) go beyond a correlation towards a causal relationship between molecular species and effects. The method is generic and can be applied to responses of any type, but is most useful for the analysis of dynamic and emergent responses such as switch points in the cell cycle. The strength of the analysis is illustrated for an existing mammalian cell cycle model focusing on the RP [Novak B, Tyson J (2004) J Theor Biol230, 563-579]. The reactions in the model with the highest RP control were those involved in: (a) the interplay between retinoblastoma protein and E2F transcription factor; (b) those synthesizing the delayed response genes and cyclin D/Cdk4 in response to growth signals; (c) the E2F-dependent cyclin E/Cdk2 synthesis reaction; as well as (d) p27 formation reactions. Nine of the 23 intermediates were shown to have a good correlation between their concentration control and RP control. Sensitivity and co-control analysis indicated that the strongest control of the RP is mediated via the cyclin E/Cdk2:p27 complex concentration. Any perturbation of the RP could be related to a change in the concentration of this complex; apparent effects of other molecular species were indirect and always worked through cyclin E/Cdk2:p27, indicating a causal relationship between this complex and the positioning of the RP.
Restriction point control of the mammalian cell cycle via the cyclin E/Cdk2:p27 complex, Conradie R, Bruggeman F J, Ciliberto A, Csikasz-Nagy A, Novak B, Westerhoff H V, Snoep J L FEB Journal, 277, 357-367PubMed ID: 20015233
 |
| Time course of the mammalian cell division cycle. A time integration for 30 h is shown for six of the intermediates of the system. The G1-, S/G2- and M-phases for one cell cycle are indicated in the graph. The RP is also depicted in the G1-phase. It should be noted that, in contrast to other switch points in the model, the RP is not a hard coded event (i.e. it is not an explicit function, but rather an emergent property of the model), and it is empirically defined as the last time point where, upon cycloheximide (CHX) treatment, the cell would not finish the division cycle it started with. The CHX treatment was mimicked in the model by reducing the translation efficiency of the ribosomes [e or Eps(t)], a parameter found in all synthesis steps of the model, from 1.0 to 0.5. This definition was taken from the original publication in which the model was described [5]. |